
清空記錄
歷史記錄
取消
清空記錄
歷史記錄

CHO細胞與293細胞高效快速表達制備克級別蛋白
20世紀80年代初亿卤,基因重組技術開始應用于動物細胞幔亥,1984年Genentech公司首次實現(xiàn)重組CHO細胞表達組織型纖溶酶原激活劑(tissue type Plasminogen Activator电谣,t-PA),標志著哺乳動物細胞表達系統(tǒng)的建立美澳。隨后竞阐,許多外源蛋白基因相繼被轉染到哺乳動物細胞,一些有價值的蛋白不斷實現(xiàn)表達硫椰,包括凝血因子繁调、促紅細胞生成素(Erythropoietin,EPO)靶草、免疫球蛋白蹄胰、尿激酶(Urokinase,UK)奕翔、乙肝表面抗原(HBsAg)和單克隆抗體等裕寨。近幾年,平均每年15種以上重組蛋白新藥通過美國食品和藥物管理局(FDA)批準派继,而且?guī)讉€重磅性的生物抗體藥專利快到期促使生物仿制藥的出現(xiàn)宾袜,全球生物抗體藥銷售額超過1200億美元/年
抗體藥物生產的宿主細胞主要NS0、HEK293驾窟、Hybridoma庆猫、 SP2/0、CHO等绅络,但CHO細胞是生產抗體藥物應用最廣的宿主細胞月培,主要有以下原因:(1)CHO細胞能夠較好的適應懸浮培養(yǎng),在生物反應器中獲得較高的密度昨稼,有利于在工業(yè)上大規(guī)模培養(yǎng)节视;(2)CHO細胞不易傳播人類的病毒[8];(3)CHO細胞能夠在無血清和化學成分明確的培養(yǎng)基培養(yǎng)假栓,它能夠確保不同批次培養(yǎng)的重現(xiàn)性寻行;(4)CHO細胞能夠在翻譯后進行修飾[9](尤其是在糖蛋白方面進行糖基化),并且其糖型與人類基本一致匾荆;(5)DHFR和GS等擴增基因能夠在CHO細胞實現(xiàn)拌蜘,并最終實現(xiàn)較高表達目的蛋白。CHO細胞類型也比較多牙丽,例如:DG44简卧、DXB11、CHO K1烤芦、CHO-S等举娩。上世紀八九十年代開始,工業(yè)上使用較早的是DHFR體系(二氫葉酸還原酶缺陷型)DG44細胞。當細胞培養(yǎng)基內還有MTX(甲氨蝶呤)時铜涉,二氫葉酸還原酶被抑制智玻,通過反饋調節(jié),使得該基因自我擴增芙代,連帶齊上下100-1000kb的基因都會擴增吊奢,如此目的基因也得到擴增,即可以提高目的蛋白的表達量∥婆耄現(xiàn)在很多單抗生產的體系依然是DG44页滚。GS體系(谷氨酰胺合成酶)CHO-K1是近些年發(fā)展的一種基因擴增篩選系統(tǒng),較DHFR系統(tǒng)有明顯的優(yōu)越性铺呵,目前在國際上得到了廣泛地認可裹驰。其原理是GS在ATP水解提供能量的同時,利用細胞內的氨和谷氨酸合成谷氨酰胺片挂。在缺乏外源的谷氨酰胺培養(yǎng)基中加入GS抑制劑甲硫氨酸亞砜亞銨(MSX)邦马,可使GS基因及與之相連的目的基因得到有效擴增,從而達到提高目的基因表達水平的目的宴卖。該系統(tǒng)的優(yōu)點主要在[16]:該系統(tǒng)不需要基因缺陷型的CHO-K1細胞株做為宿主細胞;CHO-K1細胞易于培養(yǎng)邻悬,更強壯症昏;在培養(yǎng)基中無需加谷氨酰胺,能夠避免谷氨酰胺分解造成培養(yǎng)體系中氨水平高的問題父丰,降低了工藝控制的難度肝谭,并且可有效提高細胞發(fā)酵密度和延長細胞生存時間。
適合于CHO細胞表達的Vector的基本元素包括:啟動子(Promoter)蛾扇、poly A signal攘烛、篩選標記(Selectable marker)、克隆位點(Cloning site)復制起始位點(Origin of replication)镀首,有些有報告基因(Reporter gene)坟漱。啟動子一般用的較多的是SV40和CMV兩種,而CMV啟動子一般會在目的基因(輕鏈和重鏈)的前面更哄,而SV40一般會在篩選標記的前面芋齿。篩選標記一般有兩種:代謝型和抗生素類型(或者稱擴增型基因篩選標記和非擴增型基因篩選標記)。常見的篩選標記見表1成翩,工業(yè)界用的較多篩選標記為DHFR觅捆、GS、G418和puromycin等麻敌。
表一-哺乳動物細胞表達載體中常用的篩選標記
篩選標記 | 篩選試劑 | |
代謝型篩選標記(擴增型基因篩選標記) | 二氫葉酸還原酶(DHFR) | 甲氨蝶呤(MTX) |
谷氨酰胺合成酶(GS) | 氨基亞砜蛋氨酸(MSX) | |
抗生素篩選標記(非擴增型基因篩選標記) | 嘌呤霉素抗性(Puromycin acetyltransferase) | 嘌呤霉素(Puromycin) |
殺稻瘟霉素脫氨酶(Blasticidin deaminase ) | 殺稻瘟霉素(Blasticidin ) | |
組氨醇脫氫酶(Histidinol dehydrogenase) | 組氨醇(Histidinol) | |
潮霉素磷酸轉移酶(Hygromycin phosphotransferase) | 潮霉素(Hygromycin) | |
氯林可霉素耐藥基因(Zeocin resistance gene) | 氯林可霉素(Zeocin) | |
博來霉素耐藥基因(Bleomycin resistance gene) | 博萊霉素(Bleomycin) | |
氨基糖苷磷酸轉移酶(Aminoglycoside phosphotransferase) | 新霉素(Neomycin or G418) |
轉染類型一般有穩(wěn)定轉染和瞬時轉染兩種栅炒。在單抗細胞株篩選穩(wěn)轉前通常通過瞬時轉染得到一些抗體,進行質量分析,初步判斷是否滿足我們所需要的抗體赢赊;或者進行一些前期的啟動子乙漓、密碼子等組件的優(yōu)化。穩(wěn)定轉染是外源DNA既可以整合到宿主染色體中域携,也可能作為一種游離體存在簇秒。在CHO或293細胞中進行穩(wěn)定轉染時,外源DNA是整合到CHO染色體中秀鞭,不會存在游離態(tài)趋观,因為一般質粒中復制起始位點針對原核生物,在真核生物沒有作用锋边,所以在篩選過程中皱坛,游離態(tài)的DNA會隨著傳代無法復制而丟失。
常用于哺乳動物細胞轉染方法:磷酸鈣法豆巨、陽離子脂質體法剩辟、陽離子聚合物法和電穿孔法,在工業(yè)界使用最多的應該是陽離子脂質體法和電穿孔法往扔。瞬時電轉可以做到1.0 g/L以上(大體積)贩猎,可以快速制備克級別蛋白抗體。
圖一-CHO細胞大體積電轉與脂質體轉染對比
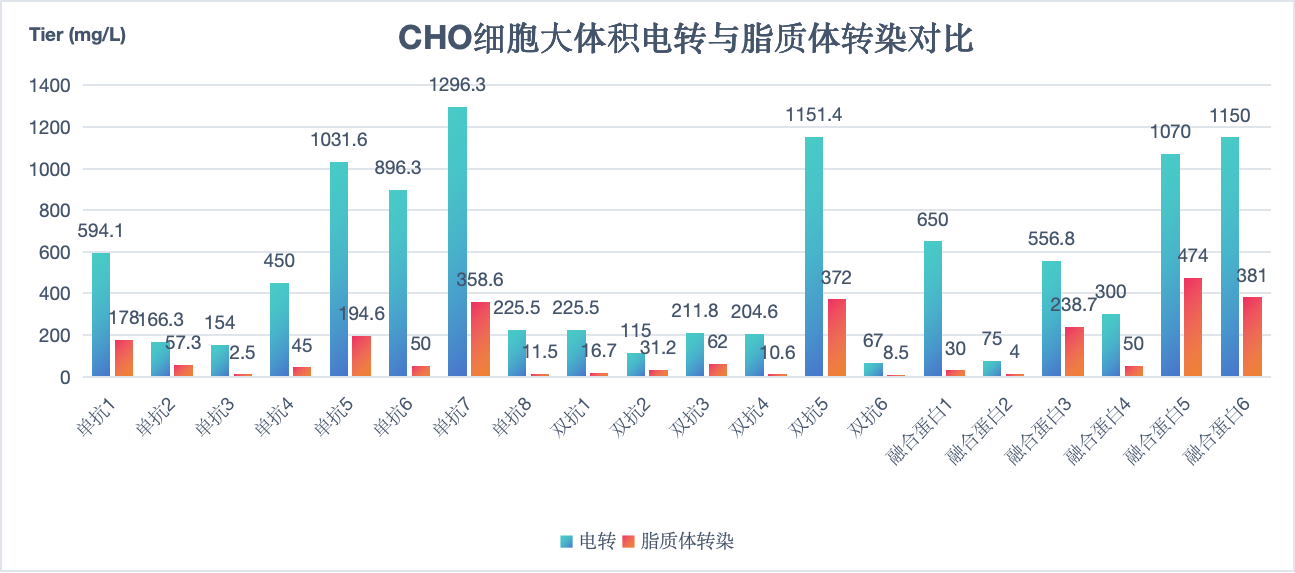
如需要制備10克或以上蛋白抗體萍膛,CHO細胞轉染之后吭服,通常會做一個pool或minipool,然后進行篩選蝗罗。結合筆者經驗艇棕,主要簡單介紹Puromycin、G418串塑、MTX以及MSX等篩選壓力原理及濃度范圍沼琉。Puromycin為氨基糖甙類抗生素,通過干擾核糖體功能阻斷哺乳動物細胞的蛋白質合成桩匪,來自鏈霉菌的pac基因具有解除puromycin毒性的作用打瘪。在篩選的時候,puromycin濃度一般在10ug/ml-50ug/ml吸祟。G418是一種氨基糖苷類抗生素瑟慈,是穩(wěn)定轉染最常用的抗性篩選試劑之一。G418在篩選的時候一般濃度范圍為:200ug/ml-700ug/ml屋匕。MTX為葉酸拮抗劑葛碧,在細胞內經過轉換后可抑制 DHFR的活性,抑制核酸合成过吻,引起細胞毒性进泼。文獻報道稱[13]蔗衡,隨著MTX濃度的增加,絕大多數(shù)細胞死亡乳绕,但在極少數(shù)幸存下來的抗性細胞中绞惦,DHFR基因得以擴增,目的基因拷貝數(shù)隨著增加洋措,提高表達量济蝉。MTX篩選時候,一般濃度范圍:25nM-1000nM菠发。MSX篩選用的是谷氨酰胺合成酶基因GS系統(tǒng)壓力王滤。谷氨酰胺合成酶(GS)擴增系統(tǒng)是新近發(fā)展的更有效的系統(tǒng),具有更高的擴增效應滓鸠。MSX篩選時候雁乡,一般濃度范圍:25uM-500uM。
圖二-CHO細胞穩(wěn)轉minipool的表達量情況
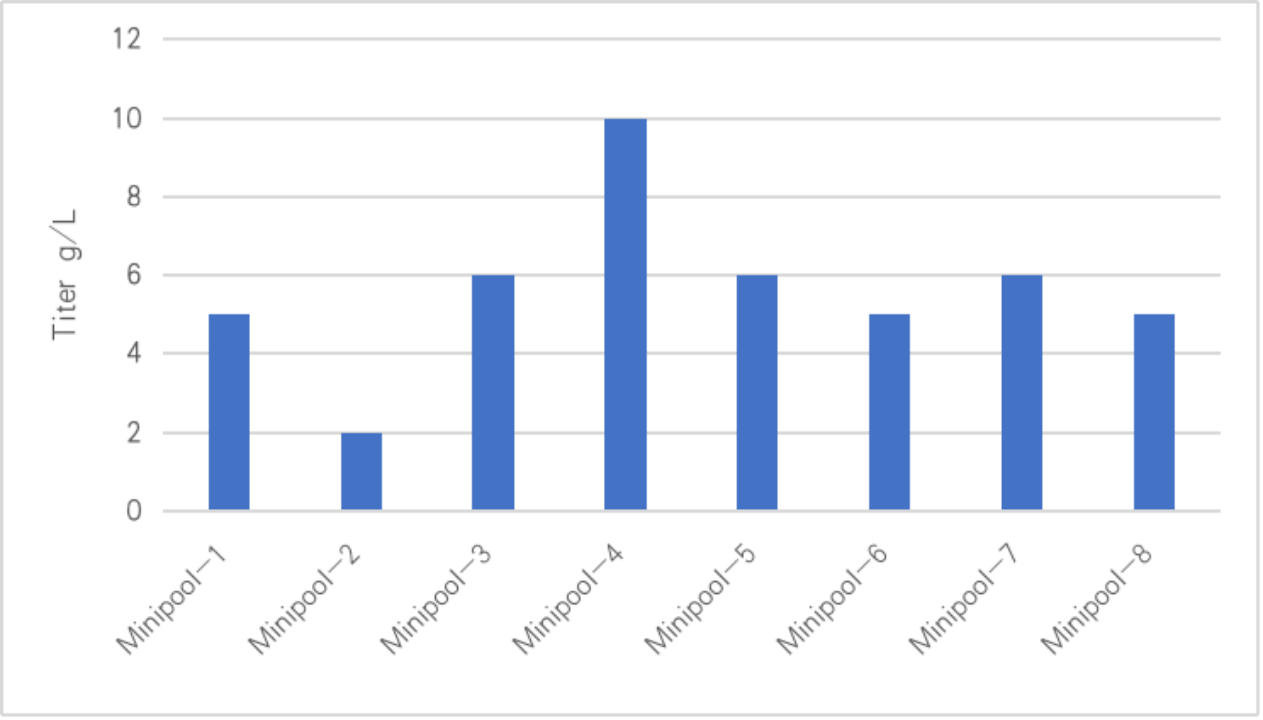
綜上糜俗,對于早研或成藥性評價過程中踱稍,需要快速制備大量蛋白抗體,有兩種較為合適的方法:1)瞬時電轉可以做到1.0 g/L以上(大體積)悠抹,可以快速制備克級別蛋白抗體珠月,周期2-3周;2)CHO細胞穩(wěn)轉之后楔敌,通常會做一個pool或minipool桥温,然后進行篩選高表達minipool,周期在6周左右梁丘。
我們的服務亮點:
24小時抗體人源化與工程改造一步到位;
3周快速表達制備克級別蛋白抗體旺韭;
極致的性價比氛谜,超預期的客戶體驗;
引用文獻資料:
[1] Kaufman, R.J.; Sharp, P.A.Amplification and expression of sequences cotransfected with a modular dihydrofolate reductase complementary dna gene. J. Mol. Biol. 1982, 159, 601–621.
[2] Wigler, M.; Perucho, M.; Kurtz, D.; Dana, S.; Pellicer, A.; Axel, R.; Silverstein, S. Proc. Natl. Acad.Transformation of mammalian cells with an amplifiable dominant-acting gene. Sci. USA 1980, 77, 3567–3570.
[3] Urlaub, G.; Chasin, L.A. Proc. Natl. Acad.Isolation of Chinese hamster cell mutants deficient in dihydrofolate reductase activity. Sci. USA 1980, 77, 4216–4220.
[4] Schimke, R.T. Gene amplification in cultured animal cells.Cell 1984, 37, 705–713. Pharmaceuticals 2013, 6
[5] Liu, P.Q.; Chan, E.M.; Cost, G.J.; Zhang, L.; Wang, J.; Miller, J.C.; Guschin, D.Y.; Reik, A.; Holmes, M.C.; Mott, J.E.; et al.Generation of a triple-gene knockout mammalian cell line using engineered zinc-finger nucleases. Biotechnol. Bioeng. 2010, 106, 97–105.
[6] Lonza launches next generation GS gene expression system. Available online: http://www.lonza.com/about-lonza/media-center/news/2012/120710-GS-System-e.aspx (Accessed on 15 February 2013).
[7]我國抗體藥物產業(yè)發(fā)展的策略探討.中國生物工程雜志 2006,26(10):93-97. LIU Yan-jun, ZHAO Yang, WANG Zheng.Strategy Analysis of Antibody Industrialization in China.China Biotechnology, 2006,26(10):93-97.
[8] Feng Li,Natarajan Vijayasankaran, Amy (Yijuan) Shen,Robert Kiss and Ashraf Amanullah, Cell culture processes for monoclonal antibody production.2010 Landes Bioscience
[8]我國單克隆抗體藥物產業(yè)化進展淺談.[J].生物產業(yè)技術,2019,(第2期).
[9] CHO細胞株開發(fā)技術策略探討. [J]. 生物技術進展. 2014, (第6期).
 瀏覽器自帶分享功能也很好用哦~
瀏覽器自帶分享功能也很好用哦~相關新聞
15502152072
( 周一至周五 9:00 - 18:00 )?

掃一掃
